Slides For This Exercise: 8, 8A, 30, 103, 3-61, 3-88, VM47, VM48, VM50
THERE IS A PRE-RECORDED LECTURE FOR THIS EXERCISE
Please go to the listing page for these to download and view it
Arteries and Veins
Let us now leave the pumping station and examine some of the pipes: the arteries and veins. Remember the definition: arteries pump blood away from the heart; veins carry blood towards the heart. No other criteria for classification of a vessel as one or the other exists. Capillaries are those small vessels within the tissues from which oxygen transfer takes place.
Arteries may be subclassified by type. Conducting or elastic arteries are large ones, with very strong and relatively elastic walls, whose function is to "conduct" the bulk of the blood to regions of the body where it's to be distributed. Examples include the aorta, subclavian, and pulmonary arteries. Once the blood has reached the region of distribution—say, the limbs—it will be handled by smaller (but still fairly large) distributing or muscular arteries, which send it to sub-regions. As the distribution area gets more and more limited the arteries become smaller. In very local areas you will see small arterioles, essentially mini-arteries with a wall considerably less muscular than the larger ones "upstream."
Elastic or Conducting Arteries
Elastic arteries are constructed like fire hoses. Rightly so, because they have the same function: to carry a stream of liquid under high pressure. Hence they're designed to minimize internal friction and flow resistance; and to maximize the strength of the wall.
The lumen is lined with a thin squamous epithelial layer (the tunica intima) which may be likened to the rubber bore of the hose; like the rubber, it offers a smooth and unimpeded passage for the flow of blood. The tunica media is a region of elastic and collagen fibers. Elastic arteries (the aorta most of all) must withstand an enormous head of pressure to pump against the peripheral systemic resistance: consequently the wall is heavily reinforced to prevent bursting, just as the wall of a fire hose has reinforcing cords in it. The elastic fibers allow some stretching and "springiness" in response to the pressure, and the collagen fibers limit the degree of stretch permitted. (In some disease or deficient nutritional states—for example lathyrism, a copper deficiency caused by certain plants—the wall may be weakened, resulting in an aneurysm which may leak, or burst with fatal effects.
To complete the analogy, the collagenous tunica adventitia of the aorta is the fabric covering on the outside of the hose. Just as the fireman needs a firm grip to control the hose, so must elastic arteries be anchored down to the surrounding structures, to prevent them from moving around as pressure varies internally. The tunica adventitia of conducting arteries is scanty, and collagenous in nature.
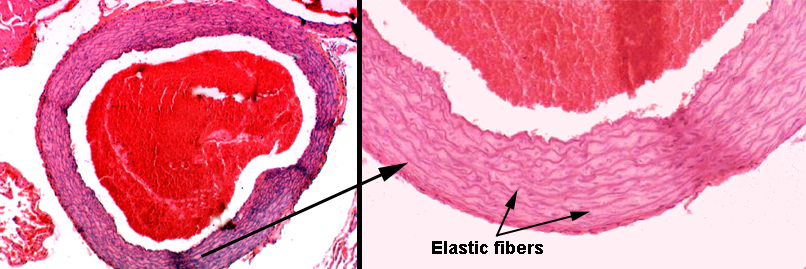
Slide 30 is a fine example of an elastic or conducting artery. It is, in fact, a section of the aorta. The section on this slide is stained with the Verhoeff's stain for elastic fibers. It renders these fibers black, and you will see that the wall of this vessel is shot through with elastic fibers. The spaces between the elastic fibers are mostly occupied with collagen, and some small amount of smooth muscle (see below). The amount of elastic fiber infiltration is so great that no internal or external elastic laminae can be identified.
This is a section of the aorta, a nice example of an elastic artery. The elastic fibers normally aren't easily seen in an H&E preparation like this one, but in this case, the reinforcement is so heavy that the concentric layers of elastic CT show up as distinct iridescent rings, some of which are marked by arrows. There's a large clot of blood in the field. The bore of this vessel—like the rest of the cardiovascular system—is lined with a simple squamous epithelium.
The innermost of the numerous elastic layers in this artery has its own name: the internal elastic lamina. It's the one right up against the tunica intima. The internal elastic lamina is much more easily seen in muscular arteries (see below) than in the elastic ones, though. In an elastic artery like this one it gets lost in the "background" of dozens of similar layers.
As is true of most elastic arteries, the tunica adventitia on this one is a relatively small contributor to the wall's thickness.
There's more than elastic fibers in the wall of this type of artery. Between the elastic layers there is a considerable amount of collagen and some smooth muscle, permitting the artery to expand under pressure and recoil to original diameter when the pressure drops again. Collagenous components in the wall prevent over-expansion and resist bursting of the vessel.
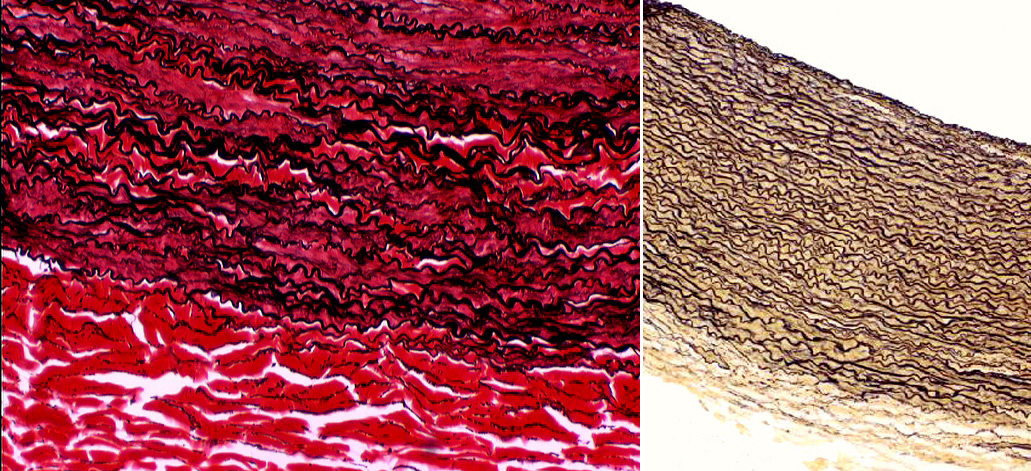
Using stains specific for elastic fibers reveals how extensive the reinforcement of the wall can be. Here are two examples, again from the aorta. The one at left has been stained with a combination of the Verhoeff and the Van Gieson stain; at about 400x, the extent of the elastic component (in black) and the collagenous reinforcement between the elastic fibers is easily visible. This field is from the outer edge of the tunica media: the almost-completely red area at the bottom is the tunica adventitia. At right, the Verhoeff stain has been used and at about 200x it clearly show elastic fibers but not the collagen or smooth muscle, which is unstained by this method.
Very large elastic arteries have their own internal blood circulation system and nervous supply. They have to: the fibroblasts and other cells that keep the wall in good shape are so far from the blood supply that diffusion won't serve their needs, and the density of the wall and its fibrillar components also impede diffusion. The smooth muscle in the wall is innervated so that the CNS can control blood pressure and initiate contractions when needed. The vasa vasorum (and in the case of nerve fibers, the nervi vasorum) i.e., the "vessels of the vessels" and the "nerves of the vessels" are a constant feature of big vessels.
Muscular Arteries
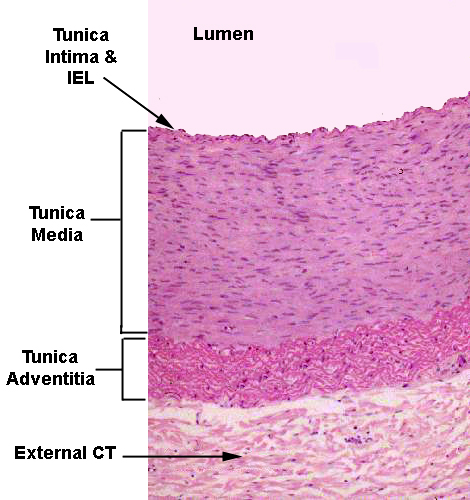 Once you get the blood out to the major regions of the body, there's a transition in the structure of the arterial wall. The proportion of elastic fibers decreases, and the proportion of smooth muscle increases. Some elastic fibers and collagen fibers will always be present, but eventually the great bulk of the tunica media will be smooth muscle, and at that point we're dealing with muscular or distributing arteries, whose function is to "distribute" blood supply to their regions of responsibility, such as a limb. The artery on slide 103, at right, is a dandy example of what a muscular or distributing artery should look like. This is the femoral artery; the femoral vein (see below) and the femoral nerve are there as well. There may be a branch point off this artery on your slide.
Once you get the blood out to the major regions of the body, there's a transition in the structure of the arterial wall. The proportion of elastic fibers decreases, and the proportion of smooth muscle increases. Some elastic fibers and collagen fibers will always be present, but eventually the great bulk of the tunica media will be smooth muscle, and at that point we're dealing with muscular or distributing arteries, whose function is to "distribute" blood supply to their regions of responsibility, such as a limb. The artery on slide 103, at right, is a dandy example of what a muscular or distributing artery should look like. This is the femoral artery; the femoral vein (see below) and the femoral nerve are there as well. There may be a branch point off this artery on your slide.
Note that the wall of the artery is mostly tunica media, and that this tunic is almost entirely smooth muscle. Elastic CT is present, to be sure, but not nearly to the extent that it is in the elastic arteries. There is also a noticeable internal elastic lamina, stained bright pink, and a less well defined external elastic lamina, though at the low magnification of this image it's hard to make out. The endothelium of the tunica intima is easily visible. The tunica adventitia grades off into the surrounding connective tissue, but is fairly sharply defined. The internal elastic lamina is just barely visible in this image. It's the undulating pink line immediately below the lining endothelium. It marks the innermost limit of the muscular tunica media. The external elastic lamina is less regular and can't be made out at all. Note the prominent collagenous tunica adventitia here. In muscular arteries the tunica adventitia is often most of the wall's overall thickness. The external CT investment anchors this artery to the surrounding CT.
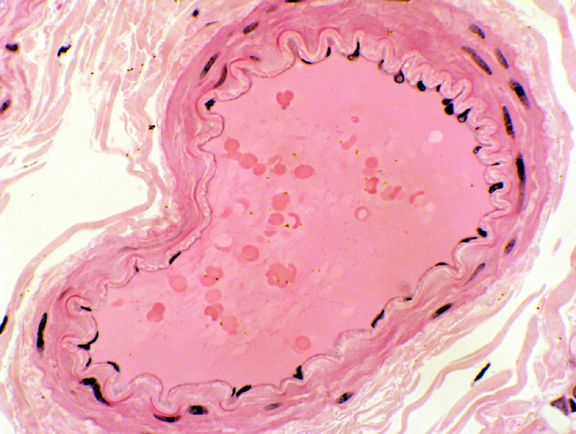 The internal elastic lamina is most easily seen in smaller muscular arteries. In these it usually stands out as a bright pink undulating band just below the lining epithelial cells and their supporting CT. In life, the artery is always under some pressure, but when death occurs the tonus of the wall causes a partial collapse, so that the undulation is something of an artifact.
The internal elastic lamina is most easily seen in smaller muscular arteries. In these it usually stands out as a bright pink undulating band just below the lining epithelial cells and their supporting CT. In life, the artery is always under some pressure, but when death occurs the tonus of the wall causes a partial collapse, so that the undulation is something of an artifact.
In the example at left, the IEL is indicated; this example also clearly shows the separation of the lining epithelium from the IEL by the sub-endothelial CT. A few layers of smooth muscle constitute the tunica media in this small artery.
 The smooth muscle of the wall of distributing arteries makes them very extensible, and also provides for a counter force to be exerted against the pressure of filling. As the vessel expands the smooth muscle cells are stretched: in reaction to this they begin to contract. The peak of their contraction comes at about the point where systole ends and diastole begins; thus the contraction of the arterial walls dampens out the pulsations of the flow to provide a more or less steady supply of blood at normal pressure into the capillary beds. Nervous input can also control this to some extent, independent of the mechanical force of stretching. As distance from the heart increases, the force required to dampen the oscillations is less, and smaller arteries can handle it; the interval between peaks also lengthens and there is a much more uniform flow rate.
The smooth muscle of the wall of distributing arteries makes them very extensible, and also provides for a counter force to be exerted against the pressure of filling. As the vessel expands the smooth muscle cells are stretched: in reaction to this they begin to contract. The peak of their contraction comes at about the point where systole ends and diastole begins; thus the contraction of the arterial walls dampens out the pulsations of the flow to provide a more or less steady supply of blood at normal pressure into the capillary beds. Nervous input can also control this to some extent, independent of the mechanical force of stretching. As distance from the heart increases, the force required to dampen the oscillations is less, and smaller arteries can handle it; the interval between peaks also lengthens and there is a much more uniform flow rate.
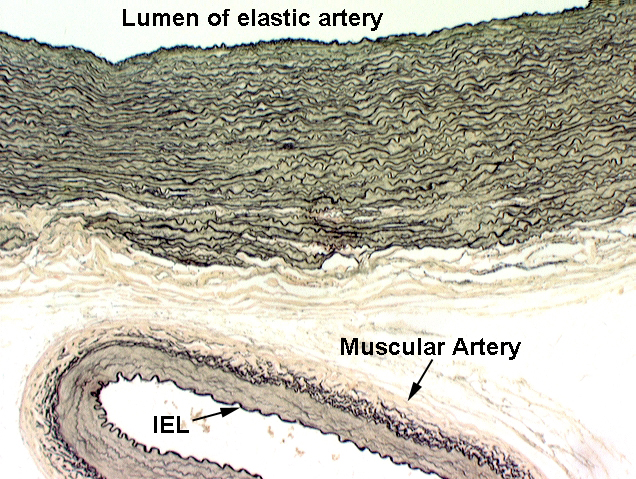
 It's useful to put the two major arterial types side by side, stained to reveal the wall components. On slide 30, this has been done using the Verhoeff stain. A small muscular artery is near the large elastic one, to make the point that as the artery size decreases, the proportion of elastic fibers in the wall decreases as well. The wall of the small artery has almost no elastic tissue in it, but the the internal elastic lamina, however, is very clearly defined against the background of the muscular tunica media.
It's useful to put the two major arterial types side by side, stained to reveal the wall components. On slide 30, this has been done using the Verhoeff stain. A small muscular artery is near the large elastic one, to make the point that as the artery size decreases, the proportion of elastic fibers in the wall decreases as well. The wall of the small artery has almost no elastic tissue in it, but the the internal elastic lamina, however, is very clearly defined against the background of the muscular tunica media.
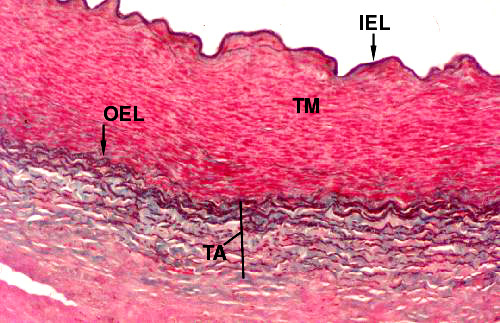
It's possible to combine several staining methods to show all the major components of the wall, including the smooth muscle. In the image below, the Verhoeff and Masson's stains have been combined. Elastic tissue is stained black; smooth muscle is red; and collagen is green. The tunica media (TM) is almost entirely muscle, with a few minor streaks of green collagen in it. The inner elastic lamina (IEL) stands out prominently; and the outer elastic lamina (OEL) demarcating the end of the tunica media is also easily visible. The tunica adventitia (TA) is a mixture of green collagen fibers and black elastic fibers interwoven with each other to provide strength and resilience.
Slide 8 demonstrates the brachial artery and vein. The artery has the typical structure of a distributing artery, and it has a very nice tunica intima and tunica media. The vein shows the typical structure of tunica intima, scanty tunica media, and a thick CT tunica adventitia. There are also valves present.
Arteries In Longitudinal Section
Since arteries are pretty common in almost any microscope slide you'll look at, it's important to be able to recognize them when they aren't cut in cross section, as all the previous examples have been.
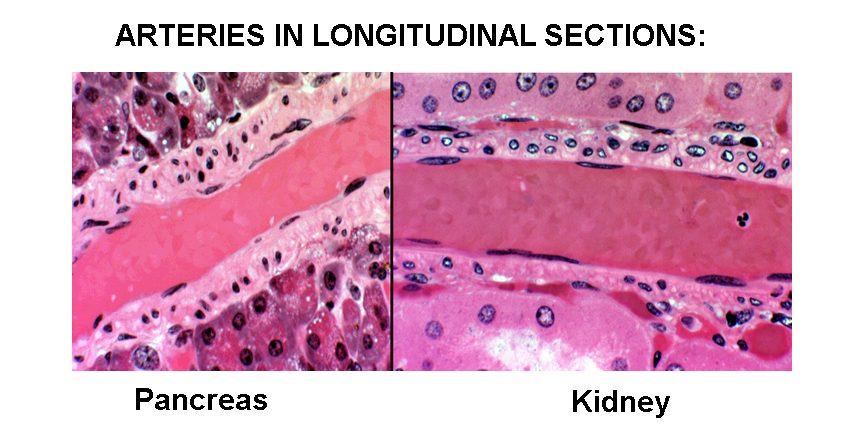
Here are two examples from tissue sections. The left section is from slide 34; the right from slide 148. Since the smooth muscle in the wall of an artery is oriented with the long axis of the smooth muscle cells around the long axis of the vessel proper, in cross sections, the muscle cells are cut in longitudinal view, but in long sections of the vessels, as here, the cells are cut in cross section. Compare these profiles (both at about 400x) to the sections of smooth muscle in Exercise 10, and you'll be able to make out the boundaries of the smooth muscle easily.
Aneurysms
Sometimes the wall of an artery—especially a big one like the aorta, which is subjected to all or nearly all the pressure the heart can generate—is weakened by disease, malnutrition, age, or some other affliction. Under a good head of pressure, blood begins to dissect the layers of reinforcing material, separating them and causing the side of the artery to bulge, exactly as a garden hose does when its wall is weakened. This bulge is an aneurysm. Chronic high blood pressure makes this more likely, though an aneurysm can occur even when systemic blood pressure is lower than normal. Needless to say, an aneurysm that bursts will really spoil your plans for the weekend. If it's a big one that blows out, such as the aorta, death will be rapid.
Veins
 Veins are those vessels leading blood back towards the heart. As a rule, they
have much thinner walls than arteries do, though in cross sectional area they're usually larger than the corresponding artery, because they have to carry the same volume of blood at a lower pressure. Since veins are on the post-capillary side
of the circulatory loop, operating at much lower pressures,
there's less need for burst
resistance. Thin walls are also important because much of the pressure that
drives blood through veins is generated not by the heart, but by contraction of
the muscles in the region of the vein. This "squishes" the blood back
through the vein. Because they have low pressures, some veins have venous valves in them
to prevent back flow. This is especially true of medium sized veins in the
extremities, as they have to lift blood against gravity.
Veins are those vessels leading blood back towards the heart. As a rule, they
have much thinner walls than arteries do, though in cross sectional area they're usually larger than the corresponding artery, because they have to carry the same volume of blood at a lower pressure. Since veins are on the post-capillary side
of the circulatory loop, operating at much lower pressures,
there's less need for burst
resistance. Thin walls are also important because much of the pressure that
drives blood through veins is generated not by the heart, but by contraction of
the muscles in the region of the vein. This "squishes" the blood back
through the vein. Because they have low pressures, some veins have venous valves in them
to prevent back flow. This is especially true of medium sized veins in the
extremities, as they have to lift blood against gravity.
(If an animal is held totally immobile for a long period of time, the return of the blood to the heart is diminished, because contraction of the muscles of the limbs no longer pushes blood back to the heart. Since the heart can only put out what it takes in, the net result of decreased venous return is decreased arterial output. At some point the output declines to the point where the brain is no longer receiving sufficient blood, and the animal loses consciousness. This happens surprisingly often. Soldiers on parade, when forced to remain at the position of "attention" for long periods, will from time to time keel over and quite literally "drop out" of the ranks as they faint.)
Venous Valves
Veins of a certain size usually have valves to prevent back flow. Veins of this type are usually found in the extremities, the valves allowing blood to move back towards the heart with less effort. Venous pressures are so low, and the propelling force has to fight against not only the resistance of the vessels but the relentless pull of gravity, that valves in the vein allow the blood to be pushed up above and when the valve closes without running back down when pressure slacks off.
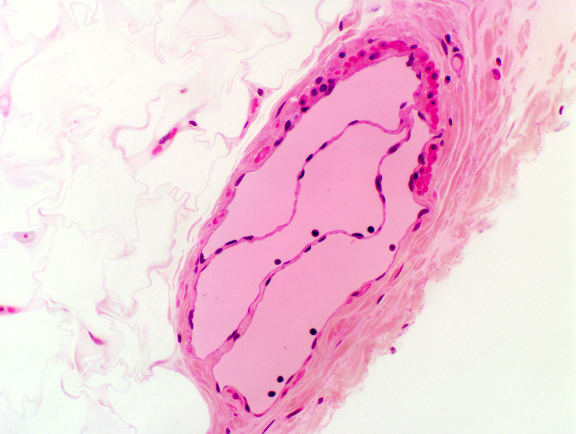 The venous valve has a great deal of structural similarity to the valves of the heart. There's a CT core with epithelium on both sides. Two valve flaps meet in the center of the vessel. You can find such valves on slide 8, and when you do, note especially the direction of the valve: it's designed to permit blood to flow in one direction only.
The venous valve has a great deal of structural similarity to the valves of the heart. There's a CT core with epithelium on both sides. Two valve flaps meet in the center of the vessel. You can find such valves on slide 8, and when you do, note especially the direction of the valve: it's designed to permit blood to flow in one direction only.
Pooling of blood occurs on the superior side of the valve flaps. In animals with long lives and vertical postures, the continual stress that the weight of this blood imposes sometimes causes outpouching of the vein. If there is a weakening of the wall on the downstream side of the valve, the blood will push it out into a small bubble analogous to the aneurysm in an artery. The result is what we call "varicose veins" that many people—especially those who work on their feet—develop in later life. This image from slide 8 shows a fairly large vein, cut more or less in cross section. The flaps on both sides of one of its valves are visible.
Capillaries are small vessels, whose walls are thin enough to allow the diffusion of nutrients, oxygen and carbon dioxide across them. They are "where the action is" in gas and nutrient exchange: with a few exceptions, no cell of the body is very far from one, because access to the blood is an absolute requirement: cell death would result from anoxia or the loss of nutrient and waste transport.
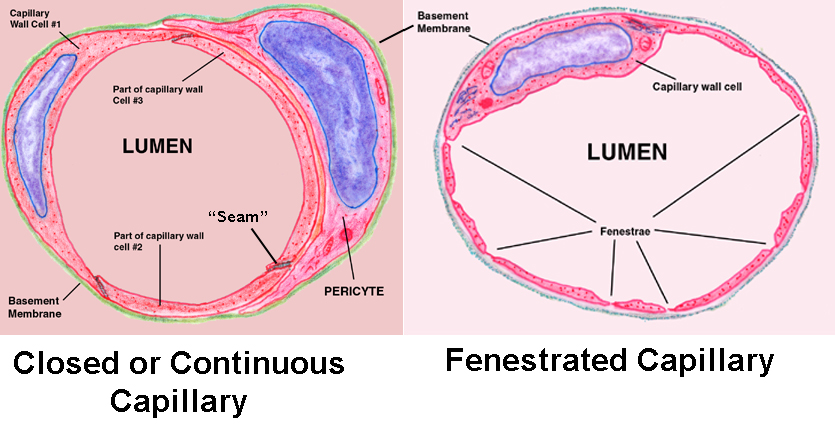
Capillaries are by far the most numerous class of vessels, though they're so small they can only be appreciated in microscopic sections. There are two types: closed or continuous capillaries, and fenestrated capillaries.
The closed type is found in locations where rapid "bulk" transfer of materials between the blood and the tissue it served isn't needed, such as in muscles. Closed capillaries can move material in and out using a process of sequential endocytosis and exocytosis, as well as simple diffusion for small molecules.
Fenestrated capillaries, which have actual pores in their walls, are located wherever immediate movement of materials is a functional necessity, such as in endocrine organs and in the kidney. They don't demonstrate the "transcytosis" activity of the closed type, because bulk movement of ions, hormones, nutrients, etc. through the pores is ample
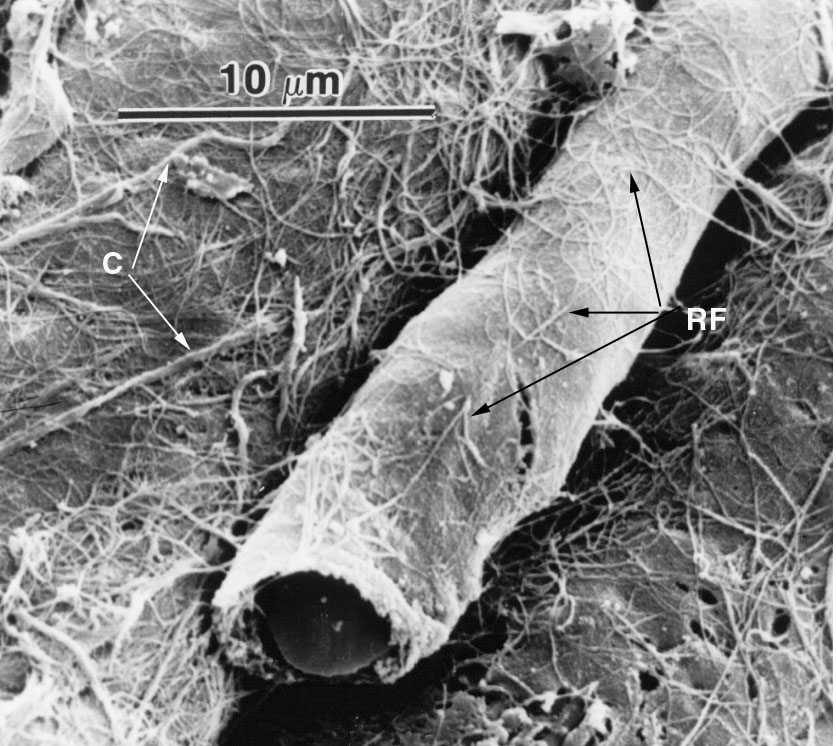 Here's a scanning electron micrograph of a capillary running alongside muscle fibers. This is a closed capillary. This striking image shows the approximate size of a capillary very well: they're just about the same size as the erythrocytes that flow through them, maybe 10 µm in luminal diameter. In this picture you can see an erythrocyte peeking out at the broken end, and the anchoring fibrils of connective tissue that attach it to the surrounding muscle mass (RF).
Here's a scanning electron micrograph of a capillary running alongside muscle fibers. This is a closed capillary. This striking image shows the approximate size of a capillary very well: they're just about the same size as the erythrocytes that flow through them, maybe 10 µm in luminal diameter. In this picture you can see an erythrocyte peeking out at the broken end, and the anchoring fibrils of connective tissue that attach it to the surrounding muscle mass (RF).
Compare this to the diagram above: the wall is made of very thin squamous cells which are sealed together at the edges by desmosomes; in other words, there's only a tunica intima, and not much of that beyond the lining epithelium and the basement membrane it rests on. In a section it's normal to see parts of several of the mural cells in the same plane, since there's quite a bit of interdigitation between the irregular borders of adjacent cells.
You may also see a second cell type, the pericyte. Technically this isn't part of the capillary wall. Pericytes "embrace" the wall but never make contact with the blood, as the mural cells do; they're believed to have some sort of contractile function. They're so closely associated with the mural epithelium they share a basement membrane with it.
 Despite their small size, capillaries are so numerous that they're easy to find. They often will have blood cells in them, which also makes them easy to spot. This one is from skeletal muscle on slide 14. Skeletal muscle has an abundant blood supply, and this capillary is lying between two of the large muscle cells. A couple of erythrocytes are visible, and the bore diameter of the capillary is just large enough to accommodate them (double arrow). A white blood cell (probably a neutrophil) is visible in the left end of the field. As it happens there are no capillary cell nuclei nor pericyte nuclei visible, they're out of the plane of the section; but the wall itself can be see. The cytoplasm of the mural cells is very scanty, and hence the precise limits of the cell and the CT around it are hard to make out.
Despite their small size, capillaries are so numerous that they're easy to find. They often will have blood cells in them, which also makes them easy to spot. This one is from skeletal muscle on slide 14. Skeletal muscle has an abundant blood supply, and this capillary is lying between two of the large muscle cells. A couple of erythrocytes are visible, and the bore diameter of the capillary is just large enough to accommodate them (double arrow). A white blood cell (probably a neutrophil) is visible in the left end of the field. As it happens there are no capillary cell nuclei nor pericyte nuclei visible, they're out of the plane of the section; but the wall itself can be see. The cytoplasm of the mural cells is very scanty, and hence the precise limits of the cell and the CT around it are hard to make out.
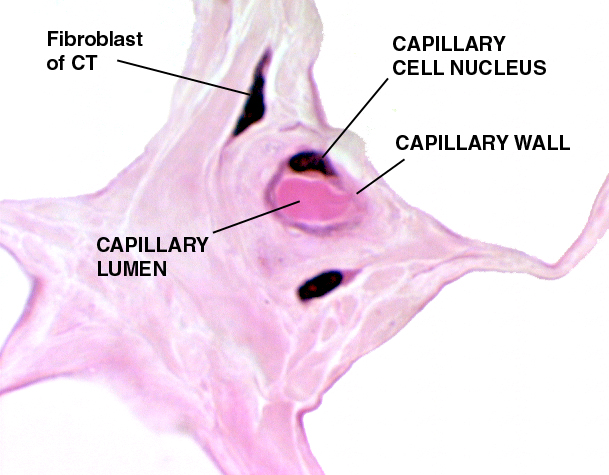 The capillary at right is from white fat. It's cut in cross section. The plane of section has passed through the nucleus of the mural cell, and the lumen is filled with the cytoplasm of an erythrocyte.
The capillary at right is from white fat. It's cut in cross section. The plane of section has passed through the nucleus of the mural cell, and the lumen is filled with the cytoplasm of an erythrocyte.
All blood vessels, of whatever type, are derived from embryonic mesoderm, the only one of the three layers of the embryo that has angiogenic potential. It shouldn't therefore be too surprising to find that connective tissues, which are also of mesodermal origin, are usually well supplied with blood vessels. That's the case here: white fat is a connective tissue, and the fibrous CT that separates each fat cell (the clear spaces in this image) from the others is a second type of CT. This capillary's function is to move the components of the fat stored in it into and out of the fat cell, and to serve the needs of this tissue for nutrient and waste transport. In young animals, which haven't had time to accumulate the "wear and tear" pigment lipofuscin in their fat cells (see Exercises 2 and 3 for a discussion of lipofuscin) the presence of so many capillaries give the fat a pinkish-white color. In albinos, animals that lack melanin, it's the blood circulating in capillaries of the eye that's the source of its pink color.
 A fenestrated capillary has "holes" in it; the word comes from fenestra, the Latin for "window." These holes are actually pores, places where the plasma membrane is perforated, to allow for the movement of bulk materials from the capillary lumen to the exterior, and vice versa. This type of capillary is found in places where the rapid movement of materials is vital to function, such as in endocrine organs, where movement of hormones into the blood is carried out. Fenestrated capillaries are also found in the kidney, again a place where quick transit between two compartments is the design criterion.
A fenestrated capillary has "holes" in it; the word comes from fenestra, the Latin for "window." These holes are actually pores, places where the plasma membrane is perforated, to allow for the movement of bulk materials from the capillary lumen to the exterior, and vice versa. This type of capillary is found in places where the rapid movement of materials is vital to function, such as in endocrine organs, where movement of hormones into the blood is carried out. Fenestrated capillaries are also found in the kidney, again a place where quick transit between two compartments is the design criterion.
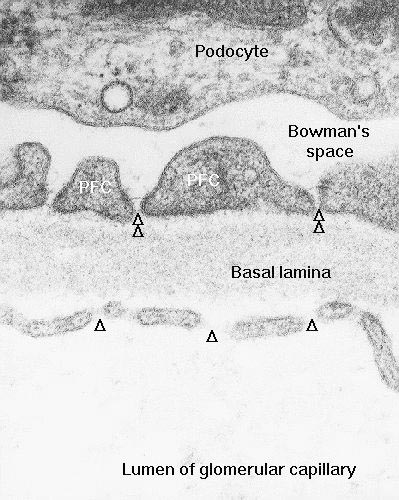
It's not possible to see the fenestrations in a light microscope preparation. To appreciate the nature of these pores, an electron microscopic image is required, because they're below the level of the light microscope's resolution. Such an image is provided here, at about 80,000 diameters. This example is from the kidney, but similar capillary profiles could be found anywhere fast movement of large molecules is important to normal function. The capillaries of the glomerulus (the tuft of blood vessels that fills the renal corpuscle) use hydrostatic pressure from the arterial supply to filter blood plasma through the pores in the formation of urine (for details, see Exercise 23). As this image makes clear, the fenestrations are actual openings in the capillary wall. The arrows show the direction of flow of materials, driven by the higher pressure in the capillary lumen than in Bowman's space. Remember, the image shown here is two-dimensional, but the cell isn't. The "breaks" indicated are really more or less circular perforations in the plasma membrane, and they have depth; they lie on both sides of the plane of the section.
Sinusoids
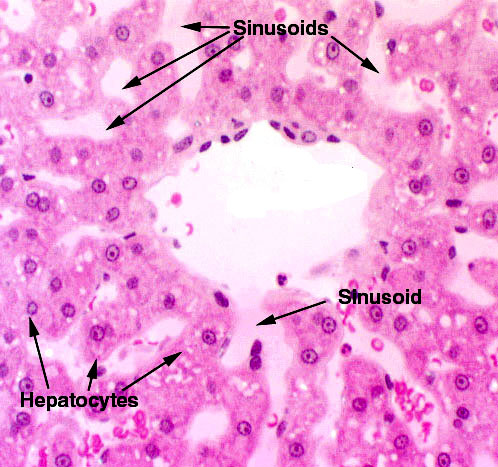 There's one more category of blood vessels to be dealt with. These are somewhat like capillaries, and might be considered as a sub-set. Sinusoids are a form of large, irregular capillary-type vessel; they have the thin, intima-only wall construction of a capillary and may be fenestrated. Sinusoids are found where slow flow, intimate contact between blood and tissue, and rapid exchange of materials are required. You'll find them on slide 123 in the liver, between plates of hepatic cells.
There's one more category of blood vessels to be dealt with. These are somewhat like capillaries, and might be considered as a sub-set. Sinusoids are a form of large, irregular capillary-type vessel; they have the thin, intima-only wall construction of a capillary and may be fenestrated. Sinusoids are found where slow flow, intimate contact between blood and tissue, and rapid exchange of materials are required. You'll find them on slide 123 in the liver, between plates of hepatic cells.
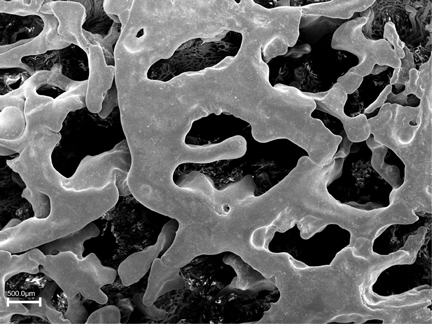 Sinusoids are flattened and irregular in shape, as the image at right makes clear. This scanning EM picture is a plastic cast of the sinusoids in the placenta of a goat, another place where slow flow and efficient transfer of materials is important to normal function. In this method the blood spaces are filled with plastic; the plastic is allowed to harden, and the tissue digested away to leave an accurate impression of the shape of the filled space.
Sinusoids are flattened and irregular in shape, as the image at right makes clear. This scanning EM picture is a plastic cast of the sinusoids in the placenta of a goat, another place where slow flow and efficient transfer of materials is important to normal function. In this method the blood spaces are filled with plastic; the plastic is allowed to harden, and the tissue digested away to leave an accurate impression of the shape of the filled space.