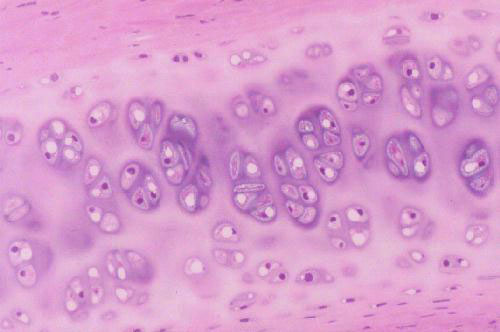
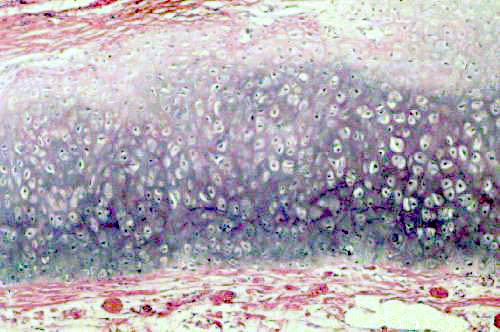
 You'll be able to find on either of these a typical field of hyaline cartilage such as the one shown at left. The cells of this tissue are chondrocytes.
You'll be able to find on either of these a typical field of hyaline cartilage such as the one shown at left. The cells of this tissue are chondrocytes. SLIDES USED IN THIS EXERCISE: 26, VM51, VM53, 75, VM05, 10, VM06
THERE IS A PRE-RECORDED LECTURE FOR THIS EXERCISE
Please go to the listing page for these to download and view it
Connective tissues are those with supportive functions. In no case is this truer than that of cartilage and bone, which are truly "supportive" in a physical (as well as physiological) sense. Cartilage forms the skeleton of mammalian embryos; in the Chondrichthyes (sharks and rays) and a few other groups a cartilaginous skeleton persists throughout life. In mammals, however, the cartilage model for the skeleton is found only in the earlier stages of development. For the most part and it's later replaced by bone, and cartilages of various kinds persist in adult mammals only in a few specific locations.
Note that word "replaced." This is an important point: cartilage is not "turned into" bone, it is replaced by bone in a complicated process which will be dealt with in Exercise 8 on bone. Not all of the cartilage of mammals is replaced. Some of it persists in areas which require resilient but flexible stiffening. Examples of persisting cartilage include the walls of larger respiratory passageways, the tip of the nose, and the ears. Cartilage is also present at the articular surfaces of joints.
There are three basic forms of cartilage: hyaline, elastic, and fibrous. Hyaline cartilage serves as the structural model, and the other two forms are described with reference to it. Many authorities regard elastic cartilage as a transitional form falling between between cartilage proper and connective tissue proper.
As with all CT's, cartilage is a combination of cells, fibers, and intercellular matrix material. The fibrillar components provide for shape retention and tensile strength, while the viscous, hydrated matrix absorbs compression forces. In the case of hyaline cartilage, the matrix material is the predominant element. In elastic cartilage, there's a lot of matrix present, but the fibrillar component differs from hyaline; and in fibrous cartilage—as you'd expect from the name—the dominant component is the fibers. We'll look at these three types in turn, starting with hyaline cartilage.
Hyaline cartilage is the most common type, found in several places. Begin with slide 1208, which includes a piece of trachea. Slide 26 is another example of trachea. This organ is supported by C-shaped rings of hyaline cartilage.
You'll be able to find on either of these a typical field of hyaline cartilage such as the one shown at left. The cells of this tissue are chondrocytes.
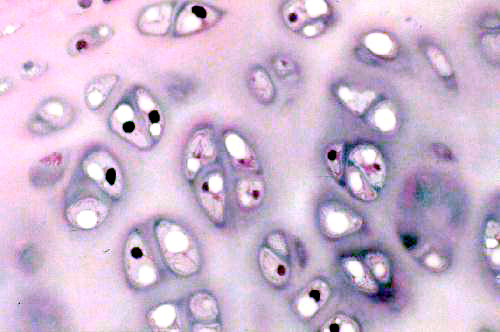
Chondrocytes individually are small cells with an oval nucleus and one or two nucleoli. The shrinkage associated with preparation makes it difficult to make out the details of these cells with the light microscope, but they have a rough surface and numerous processes that can be visualized in the electron microscope. Chondrocytes produce all the components of cartilage: the matrix material and the fibers as well. In hyaline cartilage the fibers are collagenous in nature but there aren't many of them and they aren't easily seen in an H&E preparation. As they secrete matrix material around them, the chondrocytes become walled off into small chambers or lacunae (from the Latin word lacus = "lake"). The cells are grouped into small clusters. These "cell nests" are referred to as isogenous groups. What they really represent is several generations of daughter cells that arose from the same chondroblast. In hyaline cartilage, especially in mature animals, the isogenous groups usually are somewhat separate from each other, with a perceptible amount of matrix between them. After the first chondrocyte has divided it lays down matrix between itself and the daughter cell; then the process repeats, and 3 to 5 such cells form an isogenous group.
Cartilage matrix is a homogeneous material principally composed of proteoglycans, macromolecules with a proteinaceous backbone, to which is attached complex carbohydrates (these carbohydrates are "glycosaminoglycans," usually abbreviated GAGs). The GAGs radiate from the protein core like the bristles of a bottle brush. The principal GAGs of cartilage are chondroitin sulfate and keratan sulfate. Another matrix component is hyaluronic acid, a gelatinous mucopolysaccharide. The hyaluronic acid acts as a sort of cement to bind the proteoglycans together into large aggregates.
As you might expect with this much carbohydrate present, the matrix of cartilage is very strongly positive when stained with the PAS reaction for carbohydrates, but in an H&E stained preparation it's sort of a pale pink to blue color. What little H&E stain is taken up tends to be localized into the chondrocytes, which stand out against the matrix. Notice also in the images above that there's a sort of "ring" of deeper stain close to the chondrocytes. That's where the matrix has just been formed and the density of staining is highest.
The fibrillar component of hyaline cartilage consists of a few strands of Type II collagen. Type II fibers (10 to 20 nm diameter) and unbanded. You won't be able to make them out, so don't bother to try. The matrix fills the spaces in the meshwork of Type II fibers.
An increase in the overall size, or a change in shape of a cartilaginous structure obviously has to happen somehow. Since hyaline cartilage forms the model for the bones of an developing embryo, as the embryo grows in size and develops new parts, the skeletal model has to keep pace with it. This process of growth and reshaping occurs in two ways: interstitial growth and appositional growth.
The only significant difference in the two processes is where they occur. Appositional growth takes place "at the edge" of the cartilage mass (i.e., between the cartilage proper and the surrounding perichondrium), and interstitial growth occurs "in the middle" of the mass of cartilage. As a general rule then, interstitial growth is responsible for increases in size overall, and appositional growth adds various protuberances that eventually are going to be replaced by bone to create the final shape of the structure. But the two are in essence the same thing, and completely complementary to each other. They occur simultaneously and in a completely coordinated manner.
Interstitial growth is the result of chondrocytes in their lacunae actively synthesizing matrix material. This causes the total volume of the cartilaginous structure to increase. Overall growth of the structure mainly occurs in this way.
Appositional growth occurs when the "chondrogenic" cells of the perichondrium (i.e., the cells in the overlying CT that have the potential to become chondrocytes) gear up, begin active synthesis, and start creating new cartilage at the boundaries of the structure. Eventually these new chondrocytes will become trapped in lacunae and become integrated as part of the main mass of cartilage. At that point, if they're still active in synthesis, they'll be participating in interstitial growth. By controlling the rate of interstitial versus appositional growth, the shape of the cartilaginous structure can be changed.
Cartilage doesn't usually occur "naked." With one exception it is invariably covered by CT proper. This investment is the perichondrium and it contains cells of the fibroblast line which have the potential to differentiate into chondrocytes, begin the synthesis of matrix material, and wall themselves off into lacunae.
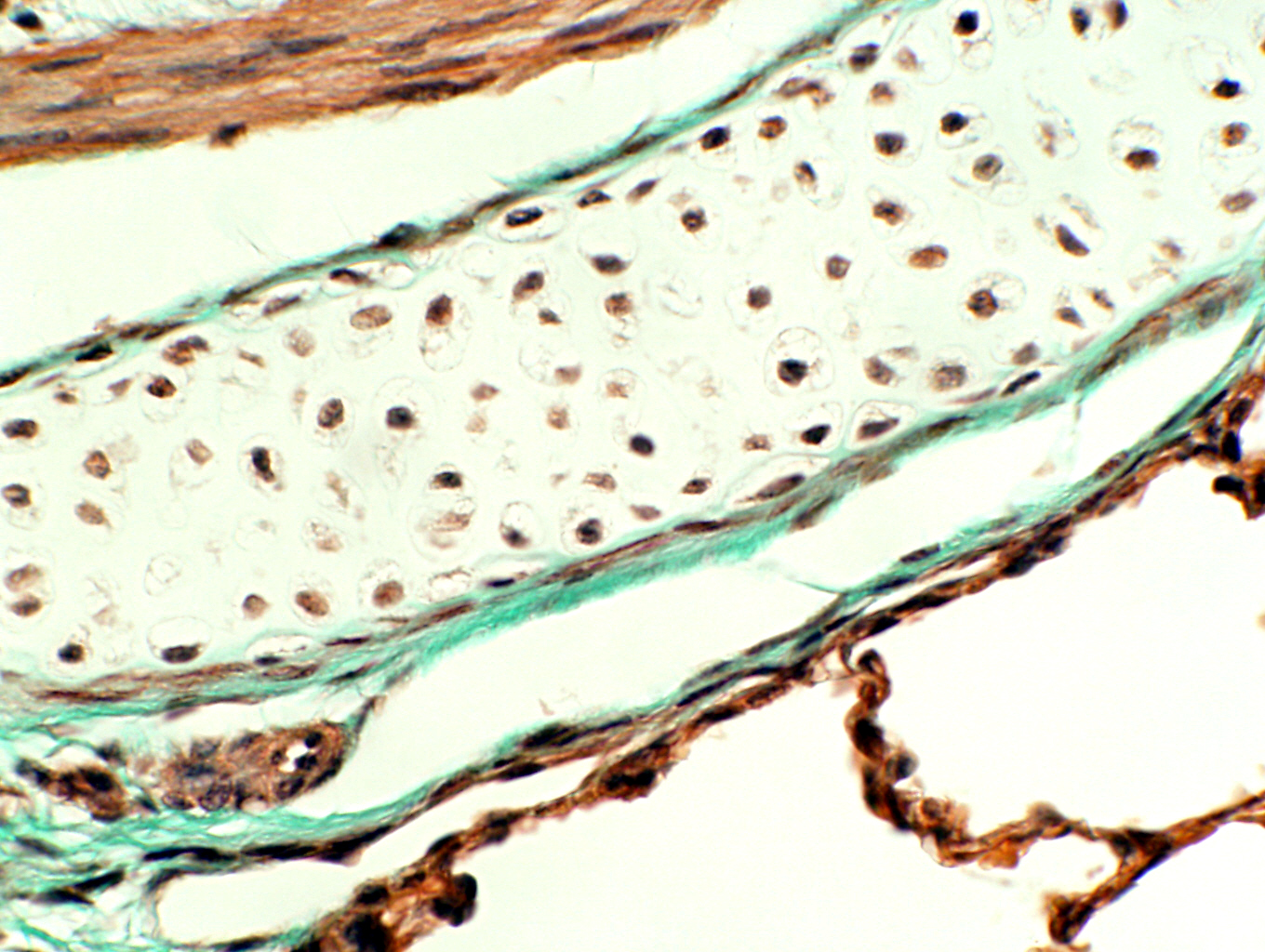 A nice example of the association of cartilage and CT proper is shown here at right. It's from slide 100. This is a piece of hyaline cartilage that's forming one of the reinforcing plates in a small airway in the lung. The slide is stained with Masson's method, which is specific for CT. The cartilage in the center is unstained, but the collagenous CT investment that covers it, the perichondrium has stained a deep green. The nuclei of fibroblasts in the perichondrium (and those of the chondrocytes in the cartilage) are brown.
A nice example of the association of cartilage and CT proper is shown here at right. It's from slide 100. This is a piece of hyaline cartilage that's forming one of the reinforcing plates in a small airway in the lung. The slide is stained with Masson's method, which is specific for CT. The cartilage in the center is unstained, but the collagenous CT investment that covers it, the perichondrium has stained a deep green. The nuclei of fibroblasts in the perichondrium (and those of the chondrocytes in the cartilage) are brown.
The exception to the rule that cartilage is always covered by a perichondrium is the articular cartilage at a synovial joint. That's what's shown at left below. The mated surfaces of this joint are covered with hyaline cartilage, and bathed in synovial fluid (which isn't visible in this slide). Hyaline cartilage is admirably adapted to use as a bearing surface. It's smooth and slick—rather like the nylon compound, "Delrin," that's used in automobile joints—so the surfaces move smoothly over each other. Wear on the surface is made up by interstitial growth in the mass of cartilage.
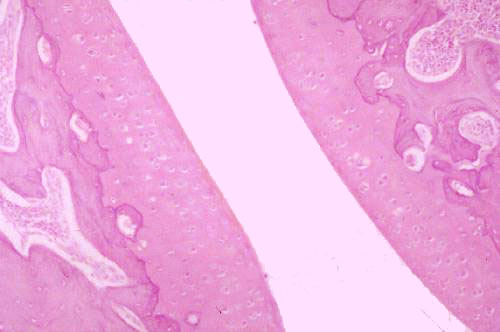 Damage to articular cartilage can be due to age, wear and tear, or abuse. Constant pounding of the joints, overworking them, or excessive weight-bearing stress can cause problems. If this happens and the cartilage degenerates to the point where bone is exposed, the joint may lose function. Untreated injuries can lead to fusion of the bone across the joint and immobility.
Damage to articular cartilage can be due to age, wear and tear, or abuse. Constant pounding of the joints, overworking them, or excessive weight-bearing stress can cause problems. If this happens and the cartilage degenerates to the point where bone is exposed, the joint may lose function. Untreated injuries can lead to fusion of the bone across the joint and immobility.
For a brief discussion of the synovial joint and its vulnerabilities, click here. Close the presentation when you're finished and return to this page.
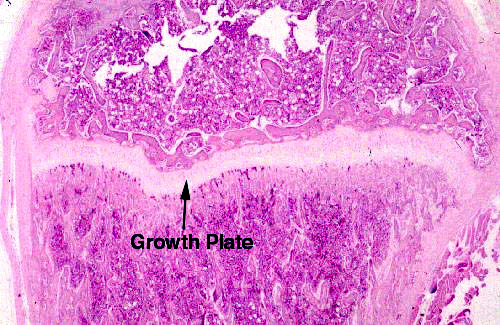 After parturition and during the growth phase of a mammal's life, the principal role of hyaline cartilage is to permit the lengthening of long bones. It does this by forming the growth plates at the proximal and distal ends of such bones. This will be dealt with in much greater detail in Exercise 8 on bone, but it can be discussed briefly here.
After parturition and during the growth phase of a mammal's life, the principal role of hyaline cartilage is to permit the lengthening of long bones. It does this by forming the growth plates at the proximal and distal ends of such bones. This will be dealt with in much greater detail in Exercise 8 on bone, but it can be discussed briefly here.
On slide 80, shown at right, you'll be able to make out a strip or band of cartilage that runs transverse to the long axis of the long bone. This is a disk of hyaline cartilage that retains its capacity for growth. The growth plates are where elongation of bones takes place (and the ONLY places where it can take place) because the cartilage can proliferate as fast as the bone formation behind it occurs, at least until maturity. Anatomically this is only one epiphysis of this bone; there's a second epiphysis at the other end.
Cartilage of the growth plates is responsive to the presence of growth hormone (GH). GH is produced in young and growing animals, so that circulating levels are fairly high. Often there's a brief period just before puberty when GH levels sharply peak, causing a "growth spurt" in adolescents. At puberty hormonal changes occur that slow down cartilaginous proliferation. Eventually the ossification process catches up and overtakes it; this is the "closure of the epiphyses" and results in their eventual obliteration. At that point no further increase in length can occur. It occurs in all mammals; the time it happens depends on the species. Dogs are skeletally mature at about a year and a half. Larger animals take longer. In humans it happens between 16 and 18 years of age.
Once the growth plates are gone, administration of exogenous bone growth stimulants (e.g., GH) causes thickening of the bones but not a lengthening. In animals whose pituitary gland is overactive (or who develop a pituitary tumor) after epiphyseal closure has occurred, this leads to a condition called "acromegaly." It's characterized by enlargement of the hands and feet, a prognathous jaw, and other skeletal deformities.
Elastic Cartilage
This form of cartilage resembles hyaline cartilage fairly closely in its structure. The principal differences between hyaline and elastic cartilage is in the nature of the fibers. It can be seen on slides 20 and 75. In elastic cartilage isogenous groups are not so well defined as in hyaline cartilage, but they still exist.
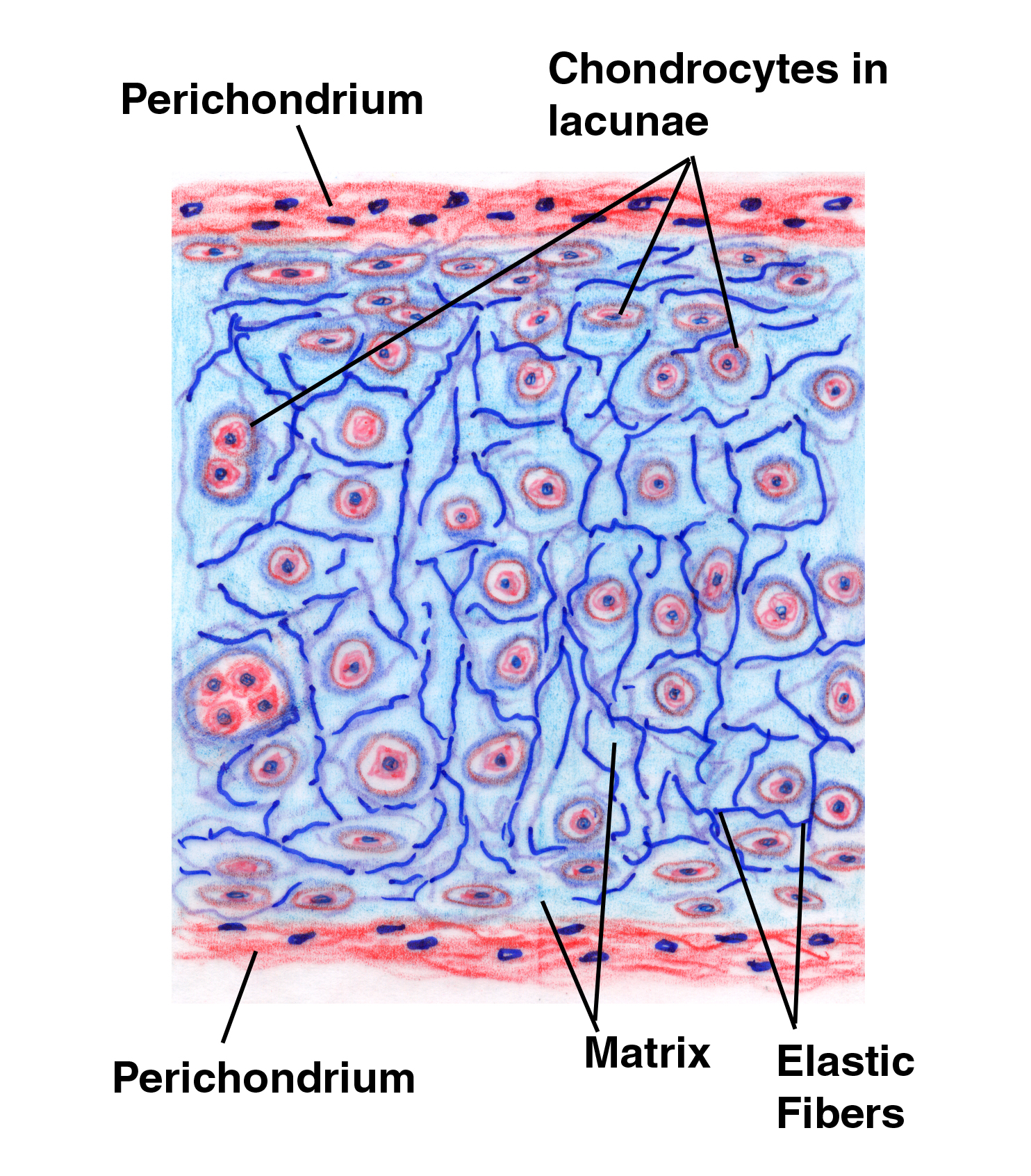
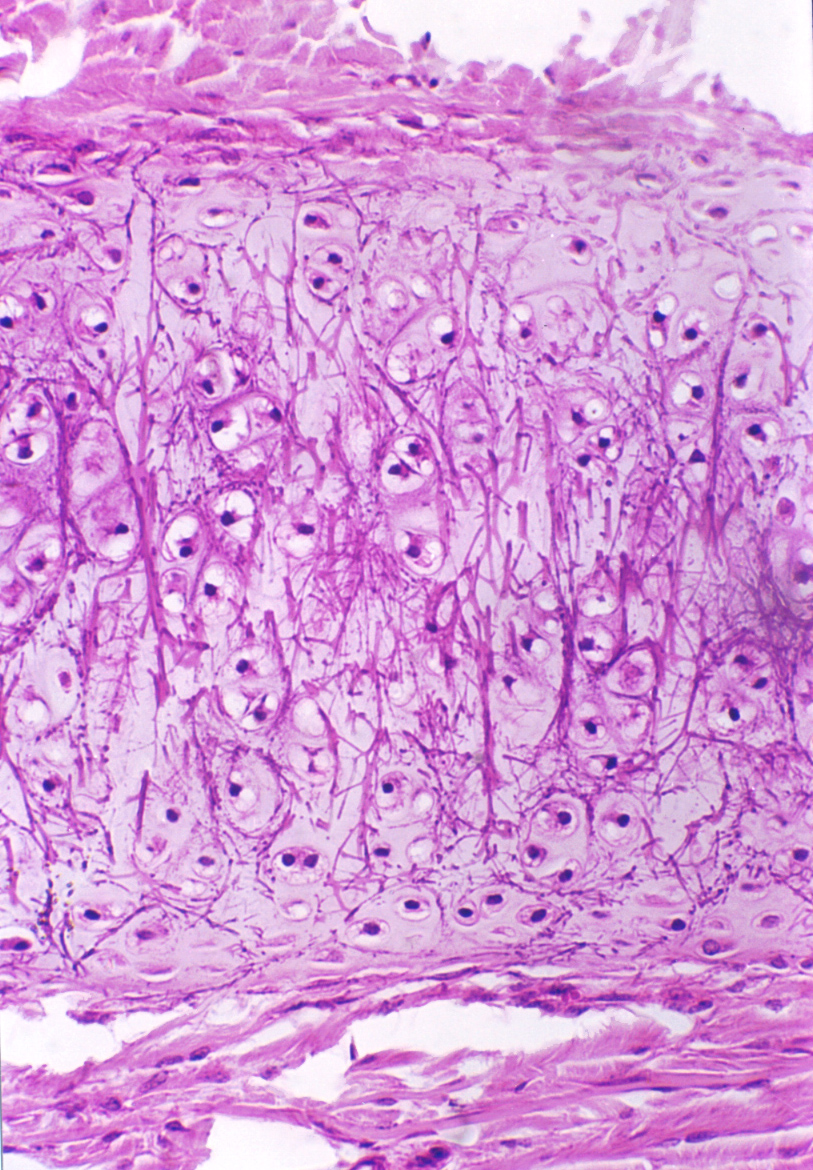
If you compare the drawing and the example shown above to the images of hyaline cartilage, it should immediately be apparent that the fibers of elastic cartilage are quite visible; they're also more numerous. The inter-group matrix material is considerably lessened in volume, and on the whole the chondrocytes of elastic cartilage are bigger than those of hyaline. The net result is that the isogenous groups are closer together and the distinct separation seen in hyaline cartilage isn't as obvious. There's still a pattern of isogenous groups but it's much harder to make them out as such. The nature of the fibrillar component also is different, in that it consists principally of elastic fibers. There's a little collagen, but the elastic fibers are predominant. They give this type of cartilage is ability to be deformed and return to shape. Notice the perichondrium on the surface of the cartilage!
 Begin with slide 20, an epiglottis. This slide is stained with orcein, a
dye which is specific for elastic fibers. In elastic cartilage, there are a
good many more fibers than there are in the hyaline type, forming a network
around and in among which the matrix material is secreted.
Begin with slide 20, an epiglottis. This slide is stained with orcein, a
dye which is specific for elastic fibers. In elastic cartilage, there are a
good many more fibers than there are in the hyaline type, forming a network
around and in among which the matrix material is secreted.
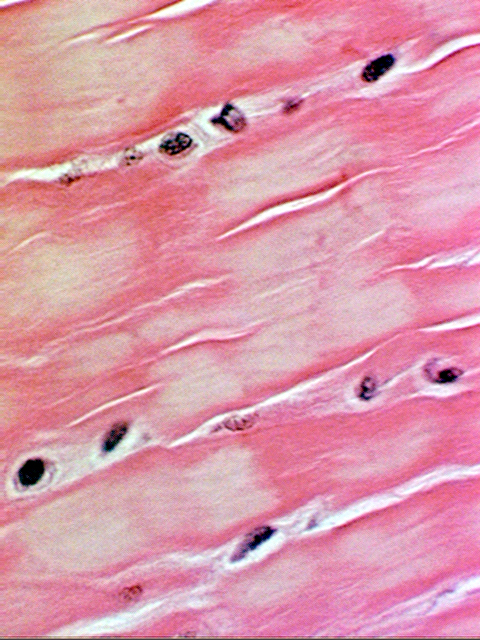
The properties of elastin and the fibers it forms give elastic cartilage its ability to be deformed and to spring back into shape immediately. Elastic cartilage is found in the pinna of the ear, the walls of the Eustachian tube, and in the epiglottis—all places in which the maintenance of a specific shape is important to proper function. The chondrocytes are more tightly packed together than is the case in hyaline cartilage, and in most histological preparations elastic cartilage tends to be more heavily stained, because of the decreased proportion of matrix material. Slide 75 provides another example. It's a section through the external ear of a rabbit. The supporting cartilage of the ear is the elastic type, and this is a nice example of the way this tissue looks in an H&E stain. Note the closely packed chondrocytes.
Fibrocartilage
The last form of cartilage is fibrocartilage. You will find it on slide 10, a section from an intervertebral disk. Fibrocartilage has only very limited distribution in the body. In addition to the intervertebral disks, it's found in the pubic symphysis, and a few other locations.
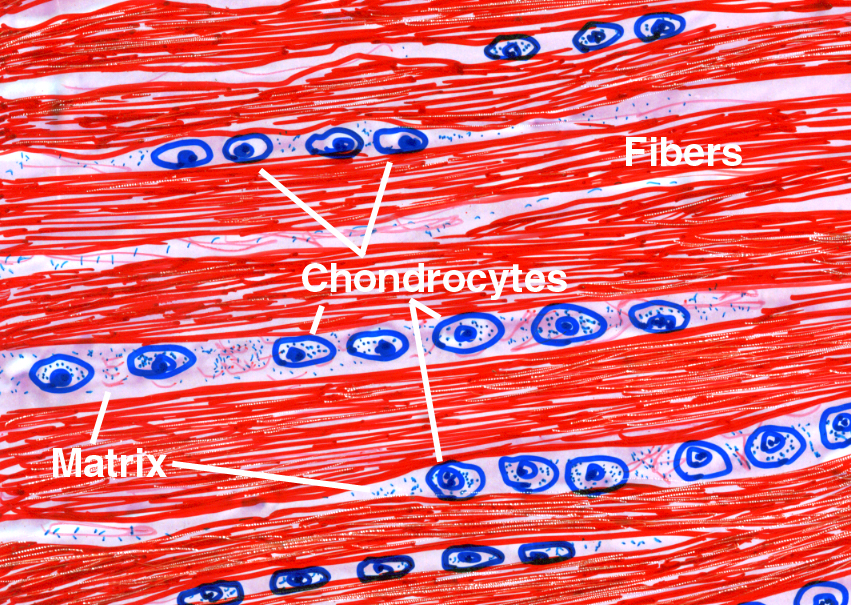
Fibrocartilage is much more similar in appearance to dense collagenous CT than to traditional cartilage, and it shares several structural similarities to both. In this type the fibrous component (which is collagen, not elastic fiber) is predominant, and the matrix is minimal. Nevertheless the cells are in lacunae, though often a lacuna may be incomplete. This is shown schematically in the sketch.
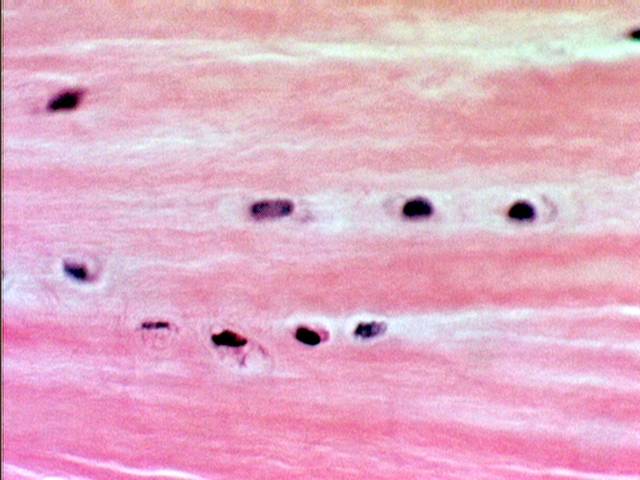 Fibrocartilage is a sort of halfway state between "true" cartilage and the fibrous CT's. The fibers are collagenous and the very scanty matrix makes it difficult to see that the chondrocytes are, in fact, located in lacunae. Because of the paucity of matrix, lacunae may be incomplete, but the cells aren't located on the fibers themselves (as fibroblasts would be in a fibrous CT). They're embedded in the matrix material between the fibers. In this higher magnification image from the same specimen you can make out the lacunae somewhat better. The presence of lacunae is the tip-off that this is, in reality, a cartilage and not dense fibrous CT.
Fibrocartilage is a sort of halfway state between "true" cartilage and the fibrous CT's. The fibers are collagenous and the very scanty matrix makes it difficult to see that the chondrocytes are, in fact, located in lacunae. Because of the paucity of matrix, lacunae may be incomplete, but the cells aren't located on the fibers themselves (as fibroblasts would be in a fibrous CT). They're embedded in the matrix material between the fibers. In this higher magnification image from the same specimen you can make out the lacunae somewhat better. The presence of lacunae is the tip-off that this is, in reality, a cartilage and not dense fibrous CT.
In fibrocartilage chondrocytes are far less numerous and much more widely separated than in other types, and as these images show, they tend to be arranged in somewhat regular rows rather than the obvious isogenous groups of hyaline or elastic cartilage.